Podcast: Play in new window
Subscribe: Apple Podcasts | Spotify | Amazon Music | Android | iHeartRadio | Blubrry | Email | TuneIn |
By Steve Williams
“I can see the fingerprints of God when I look at you I can see the fingerprints of God and I know its true you’re a masterpiece that all creation quietly applauds and you’re covered with the fingerprints of God”
~from Fingerprints of God, by Stephen Curtis Chapman
The common scientific view of the “hardware of life” (that is, the physical components of living systems) is, as Biologist Richard Dawkins puts it, “the study of complicated things that give the appearance of having been designed for a purpose.” (The Blind Watchmaker, 1986).
Unfortunately, Dawkins (like many others in his field) has succumbed to a logically fallacious assumption that a supernatural explanation is not within the “pool of live options” to explain this appearance of design. Why not? Well, to summarize the common opinions of materialists like Dawkins, it’s “not science”. But what is “Science”?
Louis Pasteur once said “Religions, philosophies, atheism, materialism, or its opposite–none of these is relevant to the matter…I might even add that, scientifically speaking, I am indifferent to them all. The question is purely one of fact”. In other words, science should be “the un-biased search for truth” without philosophical preconceptions. That definition was always the ancient understanding of the term.
Since the so-called “Enlightenment” that swept through Europe in the 1700’s, and especially since the proposal of Darwin’s Theory of Evolution in 1859, however, intellectual activists have been trying to add the qualifying concept of “within naturalistic explanations” to the definition. What that means, in effect, is the addition of a bias to the search for truth.
This bias first “got its legs” from the writings of an apparently bitter atheistic Scottish philosopher named David Hume in the 1700’s. Hume proposed a set of reasons why the supernatural should be ruled be ruled out of consideration as an explanatory mechanism. Not long afterwards, these reasons were shown to be fallacious (we’ll examine this in a later chapter), but at the time, it was as if Europe was eager to unfetter itself from religion, and atheism blossomed somewhat throughout the continent.
Most modern philosophers (even agnostic ones) find Hume’s arguments to be almost laughably illogical, but many atheists unknowingly cite him today as if he was “the Christ” of their belief system. For a full-length treatment of this subject, see John Earman’s book Hume’s Abject Failure, but suffice it to say for the moment that modern humankind has nowhere near a broad enough scope of reality to eliminate the possibility of the supernatural. To the contrary, there are many things in our experience that defy naturalistic explanations.
The philosophical name of the most common scientific form of atheism is “Materialism”, and it claims that not only is there no God, but that there is nothing even like God in the universe. Although it has taken root within the Biological sciences, it has done so to a much lesser extent within Astronomy, Philosophy, and Physics. Many Americans would probably be surprised to know that polls show that the percentage of PhD’ed scientists overall who identify themselves as Christians and who go to church is roughly the same as the percentage in the population at large. Unfortunately, the small minority who identify themselves as atheists is much louder and more aggressive though, so they exert a disproportionate influence on the media, academic standards committees and the like.
Luckily for all who respect unbiased inquiry, many Philosophers who are experts in logic by definition (logic being a subset of Philosophy), have objected vociferously, especially in the past 40 years, to this effort, and have recognized that the scientific method is at stake. As I wrote before, it seems that the key element that catalyzed this mindset since the late 1800’s is Charles Darwin’s Theory of Evolution. Biologists became so enamored with it over the years that they invested heavily in deepening and entrenching their paradigms based on that assumption, and are not willing to consider that major problems have developed within it. Pride in Biology, and a reluctance to admit being wrong might be a factor. One has to wonder if there are spiritual and carnal reasons that admitting the mere possibility of the existence of the metaphysical is so daunting to some. Since Darwinian Evolution seems to be the lynchpin of this type of thinking, let’s take a hard look at it.
The concept of life arising from non-life by random chance is called “abiogenesis”. This concept is the “creation story” of Darwinian Evolution. But what are the odds of the building blocks of life coming together by random chance in a way to provide even the possibility of life? Harold Morowitz, an agnostic Yale University physicist, created mathematical models by imagining broths of living bacteria that were superheated until all the complex chemicals were broken down into basic building blocks. After cooling the mixtures, Morowitz used physics calculations to conclude that the odds of a single bacterium reassembling by chance is one in 10100,000,000,000. (1) Wow! How can we grasp such a large statistic? Well, it’s more likely that one would win the state lottery every week for a million years by purchasing just one ticket each week!
In response to the probabilities calculated by Morowitz, Robert Shapiro, author of Origins – A Skeptic’s Guide to the Creation of Life on Earth, wrote:
The improbability involved in generating even one bacterium is so large that it reduces all considerations of time and space to nothingness. Given such odds, the time until the black holes evaporate and the space to the ends of the universe would make no difference at all. If we were to wait, we would truly be waiting for a miracle. (2)
Sir Frederick Hoyle compared the probability of life arising by chance to lining up 1050 (ten with fifty zeros after it) blind people, giving each one a scrambled Rubik’s Cube, and finding that they all solve the cube at the same moment!
Biological “Hardware” (Complex Structure) Argument
- According to a leading Darwinist, the odds of component parts in close proximity assembling into a single-celled creature are 1 in 10100,000,000,000.
- According to probability theorists, anything with lower odds than 1 in 1050is mathematically impossible.
- Therefore, the spontaneous generation of life is mathematically impossible.
Regarding the origin of life, Francis Crick, winner of the Nobel Prize in biology for his work with the DNA molecule, stated in 1982:
An honest man, armed with all the knowledge available to us now, could only state that in some sense, the origin of life appears at the moment to be almost a miracle, so many are the conditions which would have had to have been satisfied to get it going. (3)
Crick’s assessment of the hopelessness of the spontaneous generation of life on earth led him to subsequently postulate a theory called “Directed Panspermia”, which held that space aliens “seeded” life on earth. As Philip Johnson observed, “When a scientist of Crick’s caliber feels he has to invoke undetectable spaceman, it is time to consider whether the field of prebiological evolution has come to a dead end.” (4)
Ever since the discovery of DNA in 1953, the Darwinian Theory of Evolution has faced increasing challenges yearly as more and more evidence for the complexity of the cell has been discovered. In 1996, Dr. Michael Behe (professor of Biochemistry at Lehigh University) released a book entitled “Darwins’ Black Box”, which detailed an argument against Darwinian Evolution known as the “irreducible complexity” of biological structures and systems. In the 11 years since the publication of the book, it has been attacked from every angle by atheistic scientists, yet its central thesis has only gained strength, as the debate has exposed the weakness of Darwinian counter-arguments, and the naturalistic (atheistic) philosophical biases that lurk behind them.
Have you ever wondered if Charles Darwin himself would still believe in Darwinian Evolution (or macro-evolution) if he knew all of the evidence that has accumulated for and against it up to this time? Well, there is an interesting quote in which Darwin stated his own minimum standard for assessing whether or not his theory would withstand the tests of time:
If it could be demonstrated that any complex organ existed, which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down. (5)
In Darwin’s day, it was assumed that cells were very simple. In the last half of the 20th century, however, it has come to light that inside each living cell are vastly complex molecular machines made up of various protein parts. Organs, which are made up of these complex cells, have also been shown to be much more complex than previously believed.
The blueprints for assembling the protein parts for cells and organs in correct timing and order are encoded into our DNA, which is similar to binary computer code, although it is quaternary (having 4 letters instead of 2). The density of the information encoded into DNA staggers the imagination; there is enough information-storing space in a half-teaspoon of DNA to store all of the assembly instructions for every creature ever made, and room left over to include every book ever written!
In addition to the incredible information-storing capacity in DNA, there are machines and systems in biology which vastly exceed mankind’s creative capacity in terms of their complexity. For example, the blood-clotting mechanism requires a sequence of 20 different proteins (each of which has an average chance of 1 in 8.03 x 10 to the 59th power of forming by random chance!) triggering one another like dominoes falling in order, until a fibrin mesh scaffolding is formed for the clot itself.
If you subtract any one single protein (regardless of where in the sequence of 20), this scaffolding fails to form, and no blood clot is possible. Without clotting, any creature with a circulatory system would bleed to death from a tiny wound, similarly to what happens to hemophiliacs.
Now think about how this compares to Darwin’s criterion for his own theory. Macro-evolution requires a mutation for every step, each of which needs to confer an advantage in surviving or creating offspring to be retained by natural selection. Even if we grant the creation of proteins by random chance (which is extremely unlikely), at steps 1, 2 ,3, 4, etc. on up to and through step 19, there is no advantage conferred toward the production of a blood clot until step 20 is completed! If you reduce the complexity by any single component (regardless of where in the sequence the single component is), the system doesn’t work, and has no reason to be retained by natural selection. This is Irreducible Complexity.
Let’s look at another example. The Bacterial Flagellum is a tail-like protein propeller attached to one end of a bacterium that propels the organism through its environment via rapid rotations (like a miniature outboard motor driving a whip in circular motion). It has components that are remarkably similar to a man-made outboard motor, such as a rotor, a U-joint, a stator, a driveshaft, a propeller, bushings, and O-rings.
There are at least 40 different protein parts required for the assembly of a flagellum. Many of the flagellar proteins control the construction process, switching the building phases on and off with chemical triggers at just the right times, and setting up construction in the proper sequence. It is an engineering marvel. If you deduct 1% of the parts, you don’t have a 99% functional bacterial flagellum; it becomes completely dysfunctional, and you have nothing but a hindrance (probably fatal) to any organism attached to it. The following picture hints at its complexity…
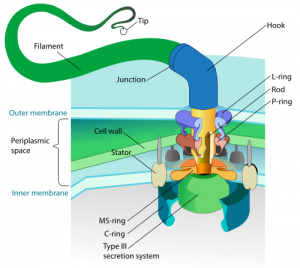
Consider the fact that it is conservatively calculated that the odds of this incredible structure forming by random chance is 1 in 10 to the 1170th power. (6) According to probability theorists, anything with a chance lower than 1 in 10 to the 50th power is mathematically impossible, so it doesn’t matter how much time you give it—it simply won’t occur by chance alone.
Just recently, a vastly more complex gear-driven, seven-engine, magnetic-guided flagellar bundle was discovered. Here is an piece on it from www.evolutionnews.org:
Souped-Up Hyperdrive Flagellum Discovered
Evolution News & Views December 3, 2012 5:05 AM
Get a load of this — a bacterium that packs a gear-driven, seven-engine, magnetic-guided flagellar bundle that gets 0 to 300 micrometers in one second, ten times faster than E. coli.
If you thought the standard bacterial flagellum made the case for intelligent design, wait till you hear the specs on MO-1, a marine bacterium described by Japanese researchers in the Proceedings of the National Academy of Sciences. Edited by Howard Berg, Harvard’s mastermind of flagellum reverse engineering, this paper describes the Ferrari of flagella.
Instead of being a simple helically wound propeller driven by a rotary motor, it is a complex organelle consisting of 7 flagella and 24 fibrils that form a tight bundle enveloped by a glycoprotein sheath…. the flagella of MO-1 must rotate individually, and yet the entire bundle functions as a unit to comprise a motility organelle.
To feel the Wow! factor, jump ahead to Figure 6 in the paper. It shows seven engines in one, arranged in a hexagonal array, stylized by the authors in a cross-sectional model that shows them all as gears interacting with 24 smaller gears between them. The flagella rotate one way, and the smaller gears rotate the opposite way to maximize torque while minimizing friction. Download the movie from the Supplemental Information page to see the gears in action.
Electron micrographs included in the paper show that the model is not unrealistic. These flagella really are tightly packed in a sheath, suggesting that the bundle acts like a gear-driven hyperdrive.
Here we have used electron cryotomography to visualize the 3D architecture of the sheathed flagella. The seven filaments are enveloped with 24 fibrils in the sheath, and their basal bodies are arranged in an intertwined hexagonal array similar to the thick and thin filaments of vertebrate skeletal muscles. This complex and exquisite architecture strongly suggests that the fibrils counter-rotate between flagella in direct contact to minimize the friction of high-speed rotation of individual flagella in the tight bundle within the sheath to enable MO-1 cells to swim at about 300 µm/s. (Emphasis added.)
At microbial level, that’s more than 10 body lengths per second. The authors were clearly excited by this engine, sounding like young men checking out high-performance cars, talking thrust, gear ratios and torque.
MO-1 is a magnetotactic bacterium capable of orienting its cell body along the geomagnetic field lines by using magnetosomes. The MO-1 cell has a flagellar apparatus with two lophotrichous [containing numerous flagella in] bundles. In contrast to peritrichously [flagella all over the cell] flagellated bacteria, MO-1 cells swim constantly in a helical trajectory toward magnetic north, and the trajectory changes from right-handed to left-handed without changes in velocity or direction. The cells are able to swim as fast as 300 μm/s, which is nearly 10-fold faster than E. coli and Salmonella. Although the flagella of the other types of bacteria usually work individually or by forming a loose bundle to produce thrust, the flagellar apparatus of MO-1 is a tight bundle of seven flagella enveloped in a sheath made of glycoproteins. This unique architecture appears to be essential for the smooth and high-speed swimming of MO-1.
They can’t see actual gears, of course, but physics demands that the mechanism of rotation must have something like it:
We hypothesize that, whereas each of the seven flagella has its torque-generating motor, the 24 fibrils counter rotate between the flagellar filaments to minimize the friction that would be generated if the flagella were directly packed together in a tight bundle. A schematic diagram representing our hypothesis is presented in Fig. 6. The flagella are represented as large brown gears and the fibrils are represented as small blue-green gears. The flagella and fibrils rotate counterclockwise and clockwise, respectively, as indicated by the arrows, to minimize friction (Movie S1). Although there is no direct evidence that the fibrils can rotate freely in the opposite direction as the flagellar filaments with which they are in direct contact, we think this is the simplest interpretation to explain the superior function afforded by the complex architecture of the MO-1 flagellar apparatus.
Considering the very tight packing of the 7 flagella and 24 fibrils that are in direct physical contact within the sheath, there appears to be no other way for the flagella to rotate at high speed without the counter rotation of the intervening fibrils. Although the fibrils and the surrounding sheath are in direct contact, the friction between them would be small because of the stocking-like flexibility of the sheath. This design must be playing an essential role in the fast, smooth rotation of the flagellar apparatus that allows the rapid swimming of MO-1.
With powerful evidence of design like this, did the researchers become converts to intelligent design? We can’t know, but would PNAS have printed such a paper without an obligatory tribute to unguided materialistic evolution? Evolution is not mentioned until the last paragraph:
Taken together, these features of the MO-1 flagellar apparatus represent an advanced level of evolution of a motility apparatus. It is also intriguing that the same pattern of an intertwined hexagonal array in two evolutionary distant systems: the basal bodies of flagella and fibrils of the MO-1 flagellar apparatus, and the thick and thin filaments in vertebrate skeletal muscle. Similar architectures of filamentous structures presumably evolved independently in prokaryotes and eukaryotes to fulfill the requirements for two very distinct mechanisms to generate motion: counter rotation and axial sliding.
OK, so the Darwinists got their offering, but it leaves a bad aftertaste: now, they have to believe that advanced mechanisms for generating motion evolved not just once, but twice — completely independent of each other. Thanks a lot, guys. Wait till the intelligent-design movement hears about this.
Oops, too late.
(http://www.evolutionnews.org/2012/12/souped-up_flage066921.html).
Going back to our previous subject, what if we just ignored the previously mentioned problems of forming the first cell, and assume that we’re starting the Darwinian process from the bacterial level and advancing to the human level? On page 153 of the book Who Was Adam?, Fazale Rana and Hugh Ross cite one of the world’s most prominent evolutionists, Dr. Francisco Ayala of UC Irvine, as calculating the minimal odds of human beings evolving from the bacterial level to be 1 in 10 to the 1 millionth power. Three physicists, John Barrow, Brandon Carter and Frank Tipler, did roughly the same calculation but included some important factors that Ayala overlooked, and came up with the number 1 in 10 to the 24 millionth power. Again, according to probability theorists, any event with lower odds than 1 in 10 to the 50th power is mathematically impossible. Therefore unguided Darwinian evolution is mathematically impossible!
Reduced to a propositional argument, it might go like this:
Biological “Hardware” (Complex Structure) Argument
- According to leading Darwinists, odds of humans evolving from a single-celled creature are 1 in 1024,000,000.
- According to probability theorists, anything with lower odds than 1 in 1050is mathematically impossible.
- Therefore, Darwinian evolution of human beings is mathematically impossible.
Now, these two sets of odds (totaling to 1 in 10100,024,000,000) seem overwhelming to say the least; why would scientists insist that creations like these could have come about by evolution? To re-iterate, it seems that biological science has become dominated by atheistic philosophers. Science is “a search for truth”, but the oligarchy in control in this day and age is trying to change that to “a search for truth by naturalistic (atheistic) means”. To them, the idea of God is unacceptable, so science cannot consider even the possibility that God created this universe and all that is in it.
Take a look at the following quote by prominent Darwinist Richard Lewontin, and consider whether his viewpoint is logically sound. Unfortunately, this quote seems to be representative of how many Darwinists think, and how they want everyone else to think:
Our willingness to accept scientific claims that are against common sense is the key to an understanding of the real struggle between science and the supernatural. We take the side of science in spite of the patent absurdity of some of its constructs, in spite of its failure to fulfill many of its extravagant promises of health and life, in spite of the tolerance of the scientific community for unsubstantiated just-so stories, because we have a prior commitment, a commitment to materialism. It is not that the methods and institutions of science somehow compel us to accept a material explanation of the phenomenal world, but, on the contrary, that we are forced by our a priori adherence to material causes to create an apparatus of investigation and a set of concepts that produce material explanations, no matter how counter-intuitive, no matter how mystifying to the uninitiated. Moreover, that materialism is absolute, for we cannot allow a Divine Foot in the door. (7)
One wonders why such a concerted effort is made to deny the metaphysical into the pool of live options. Perhaps the following quote by another prominent Darwinist named Aldous Huxley provides some insight:
I had motives for not wanting the world to have meaning; consequently assumed that it had none, and was able without any difficulty to find satisfying reasons for this assumption … For myself, as no doubt, for most of my contemporaries, the philosophy of meaninglessness was essentially an instrument of liberation. The liberation we desired was simultaneous liberation from a certain political and economic system, and liberation from a certain system of morality. We objected to the morality because it interfered with our sexual freedom. (8)
Now, just for fun, look at the following Bible passage, and think about how it relates to the quotes above:
For since the creation of the world His invisible attributes, His eternal power and divine nature, have been clearly seen, being understood through what has been made, so that they are without excuse.
For even though they knew God, they did not honor Him as God or give thanks, but they became futile in their speculations, and their foolish heart was darkened. Professing to be wise, they became fools. –Romans 1:20-22.
Perhaps this is a good place to ponder a quote from Nobel-Prize winning organic chemist Christian de Duve:
If you equate the probability of the birth of a bacteria cell to chance assembly of its atoms, eternity will not suffice to produce one… Faced with the enormous sum of lucky draws behind the success of the evolutionary game, one may legitimately wonder to what extent this success is actually written into the fabric of the universe. (9)
God has indeed left His signature in nature in its irreducible complexity and fine-tuning. Darwinism has failed repeatedly when tested as an explanation for the existence of life. Hundreds of scientists have recognized this and have signed a document called the “Dissent from Darwinism” (http://www.dissentfromdarwin.org/) to express their disagreement with philosophical naturalism dominating science through Darwinism. We simply need to “have eyes to see, and ears to hear”, and stop listening to atheistic philosophers disguised as scientists, who try to insist that the supernatural or metaphysical is off-limits for science. “Reasonable faith” is going in the same direction to which the evidence is pointing. The teachings of the Bible, understood properly, merge perfectly with science.
This article is chapter 3 From What Your Atheist Professor Doesn’t know (But Should), by Steve Williams
Original Blog Source: http://bit.ly/2Ch6MsY




